普洱茶是云南的一张亮丽的名片,也是颇具品牌价值的茶叶名品。很多爱茶的茶客或多或少都能说出几座云南的知名茶山,民间也有“易武为王、景迈为后;左相班章、右将临沧;南糯在前,布朗在后”等诸多说法。古六大茶山大都位于澜沧江沿岸,归属于西双版纳、普洱等区域。人们喜爱普洱茶蕴含着岁月千锤百炼后的滋味,更喜欢普洱茶那份经光阴沉淀后的风骨,对普洱茶的探源也从未间断。

景迈山云海记者陈飞摄
普洱茶名重天下
云南省有着悠久的茶叶生产历史和丰富的种质资源,是世界公认的茶树原产地和中国重要的茶叶生产基地。普洱茶的种植遍布云南省全境,尤其集中在澜沧江中下游地区的普洱茶区、西双版纳茶区、临沧茶区和保山茶区。普洱茶叶形较大,富含茶多酚、儿茶素和茶碱。
普洱茶的名称因族名(或因地名)而得。至唐朝,普洱茶开始了大规模的种植生产,称为“普茶”;宋明时期,普洱茶被中原逐渐认识,并且在国家社会经济贸易中开始扮演重要的角色。到了清朝(1636年—1912年),普洱茶到达第一个鼎盛时期,《滇海虞衡志》称:“普茶名重天下……茶山周八百里,入山作茶者数十万人,茶客收买,运于各处”……
千百年来,普洱茶作为云南的“土特产”,生于西南边疆,长在红土高原,随着茶马古道运往各地。清朝学者阮福记载说:“普洱茶名遍天下,京师尤重之。”普洱茶既是西南地区各民族日常饮品,更是西藏、内蒙古、甘肃等地游牧民族每天不可缺少的生活必需品,并且登堂入室,成为王公贵胄杯中的佳品。唐代以来,为顺应当地人民需求,在中国西南和西北地区,以茶叶和马匹为主要交易内容,逐渐形成了以马帮为主要运输工具的商品贸易通道,即茶马古道。茶马古道以川藏道、滇藏道与青藏道(甘青道)三条大道为主线,辅以众多的支线、附线,构成的一个庞大的交通网络,地跨陕、甘、贵、川、滇、青、藏,外延达南亚、西亚、中亚和东南亚各国,成为中国西南民族经济文化交流的走廊。
数百年的光阴,茶马古道上的石板被马蹄踩踏出一个个马蹄窝,留下了历史的印记。而沿着茶马古道源源不断向外输送的普洱茶,经过一两年甚至更多时间的艰难跋涉,在风霜雨雪、日晒雨淋、马背热汗的加持下,晒青普洱茶鲜活的细胞在穿山越岭的路途中开始自然发酵,并在后期的存放过程中继续完成自然发酵过程,转化为难以复制、独具韵味的“普洱茶”。

寄生于茶树上的“螃蟹脚”记者王毅摄
茶源之争
说到普洱茶的分类学位置,不得不提一个典故。1824年,驻守印度的英军少校勃鲁士,宣称在印度阿萨姆邦(Assam)发现野生茶树。1838年,勃鲁士印发宣传册,列举他在阿萨姆发现的多株高大野生茶,并断言印度是茶树原产地。鉴于当时英国的国际影响力,在勃鲁士和英国学者的诱导下,印度被先入为主地标榜成茶源地,普洱茶的拉丁学名也成了“assamica”。有关学术争议延续到了1981年,我国植物学泰斗张宏达教授多番实地考察,发现和确认所谓“阿萨姆种”是中国原种,起源于中国西南部。张宏达教授凭藉充分的科学依据,为学界正本清源,将var.assamica中文定名为“普洱茶种”,奠定了中国作为世界茶树原产地的地位。
世界山茶属研究权威专家、中国科学院昆明植物研究所闵天禄教授在其著作《世界山茶属的研究》中对普洱茶的起源问题进行了阐释:“TheaassamicaMasters(阿萨姆茶)是根据英属东印度茶叶公司采自阿萨姆茶园栽培的标本命名发表的。参与阿萨姆茶树调查的C.Bruce指出:阿萨姆的茶树可能是当地人从远东带回来栽培的。我们翻阅了英国各大植物标本馆的标本记录,印度阿萨姆地区未见有野生普洱茶存在的确切记载。”
野生普洱茶的自然分布限于中国南部、西南部和邻近的中南半岛北部,以云南南部森林中最为常见。西南林业大学郭荫卿教授在讲授《树木分类学》时明确指出:根据植物学家对野生普洱茶分布区域的系统调查结果和国内外重要标本馆收藏的山茶属标本及其采集记录,有关普洱茶大量的标本采集和引种记录都源于澜沧江中下游的森林地带,特别是以临沧至普洱段最为集中。这一区域也是当时来自英法等西方的植物学家和传教士最重视的地方。他们在采集了这些植物标本后大都收藏在英国和法国的标本馆,普洱茶就是当年英国植物学家、传教士采集后引种到与澜沧江流域气候相近的印度阿萨姆地区的植物之一,并在阿萨姆种植发展。而英国植物学家马斯特(Masters)是在印度的阿萨姆邦的茶园中采到了普洱茶标本后命名发表。因此,普洱茶真正的原产地应为云南的澜沧江中下游地区,这一地区也是世界山茶属茶组种类分布最集中的地区,也就是普洱茶的起源地之一。
完整的证据链
生态环境专家杨宇明教授分析了普洱茶在云南起源并保存的地理学原因:澜沧江发源于青海省玉树州杂多县,从海拔5000多米的唐古拉山脉北麓的富吉山一路南下,跨越近30个纬度,在中国境内称澜沧江,出我国国境后被称为湄公河,经老挝、缅甸、泰国、柬埔寨、越南,由越南胡志明市注入南海,是北半球南北跨越距离最长,连接着青藏高原与中南半岛的第一大河。由于澜沧江流域是连接着青藏高原与中南半岛的重要生物廊道,不仅物种分布集中,而且是不同生物地理区系成分的交汇过渡地带,其中山茶科山茶属等被子植物的古老类群得以在澜沧江河谷演化发展,成为山茶属茶组的分布与分化中心。至今,澜沧江中下游仍然是山茶属植物种类分布最多最集中的河谷地区之一,目前集中分布了如大理茶、厚轴茶、德宏茶、大苞茶和普洱茶等诸多种类,普洱茶就是其中最著名的古茶种。
与此同时,发现于普洱市景谷县,距今约3520万年前的茶树化石,从学术上最终确立了茶树的发源地在云南的事实,是世界茶树演化过程中重要古植物化石证据,学术价值不可估量。云南茶区,有数不清的野生古茶树、古茶树群落,这些古茶树成为中国作为世界饮用茶树原产地的“活化石”。资料显示,我国古茶树有5600多万株,约97%分布在云南。全球茶组植物共有31个种4个变种,其中云南有23个种3个变种,以云南茶树作模式标本定名的有16个种2个变种。作为世界饮用茶树原产地,云南分布有大量野生茶树群落,是全球独一无二的古茶树王国。据云南省林业和草原局的古茶树资源调查结果显示,云南有古茶树91万亩、约5400万株,是迄今所知世界上古茶园保存面积最大、古茶树和野生茶树群落保存数量最多的地方。
西南林业大学教授,云南古茶研究中心主任蓝增全介绍,云南境内发现的野生型、过渡型和栽培型古茶树代表,为证明云南是世界茶树原产地提供了有力证据。比如,1951年在勐海县南糯山发现的栽培型古茶树,树高8.8米、基径138厘米,树龄约800年;1961年在勐海县巴达山发现的野生型古茶树,树高32米、基径100.3厘米,树龄约1700年;1991年在澜沧县邦崴村发现的过渡型古茶树,树高11.8米、基径78.9厘米,树龄1000多年。

大平掌古树茶记者陈飞摄
全球首个茶主题世界文化遗产
隐秘在临沧市双江县勐库大雪山人迹罕至的原始森林中,集中分布着面积约1.27万亩双江县勐库镇邦马大雪山野生茶树群落,海拔2200—2750米,是目前所知野生古茶种群分布海拔最高、种群密度最大的野生古茶树群落。而普洱市澜沧拉祜族自治县景迈山的万亩千年古茶园,则是世界上保存最完好、年代最久远、面积最大的与森林乔木混种的人工栽培型古茶园。国内外专家学者赞誉为“茶树自然博物馆”,是茶叶生产规模化、产业化的样本,更是世界茶文化的根和源。
2010年,针对世界文化遗产领域茶主题“空白”,国家文物局启动“普洱景迈山古茶林文化景观”申遗工作,并于2021年1月经国务院批准后正式报送联合国教科文组织。而为什么是景迈山?我们在众多解释说明文件中找到这样一些表述:“中国西南地区是全球著名的第三纪孑遗植物的庇护所,也是山茶属的分布中心和起源中心。山茶科树木在第四纪多次冰期中存活下来,适应不同自然条件而演化成今天纷繁多样的现代茶组植物。进入全新世(地质年代约12000~10000年前至今)以后,野生古树茶的使用价值被人类发现,之后逐步被驯化并大规模栽种,这一地区遂成为中国栽培型茶树起源地,并在近代传播到了欧亚大陆、非洲和南美洲。该地区的云南省澜沧江下游流域,北回归线附近,受到西南季风影响,水热条件十分优越,加之青藏高原和横断山脉阻挡了来自北方的寒冷气流,成为了全球普洱茶的故乡。公元10世纪,布朗族先民迁徙至景迈山,发现和认识野生茶树,利用森林生态系统,与后续来到的傣族等各族一起,探索出‘林下茶’种植技术,历经千年的保护与发展,形成这一林茶共生、人地和谐的独特文化景观……”
随着2023年9月17日“普洱景迈山古茶林文化景观”申遗成功,成为中国第57项世界遗产,全球首个茶主题世界文化遗产,中国云南作为野生普洱茶的起源地将无可争议地载入史册。
(吴静波 杨宇明 作者 分别系中共云南省委宣传部原副巡视员;国家濒危物种科学委员会委员)
来源:云南网,信息贵在分享,如涉及版权问题请联系删除
茶树在生长发育过程中,虽然对不利气象条件有一定的抗衡能力,但是超过一定限度后就会产生危害,轻则造成茶叶减产、品质下降,重则使茶树死亡,那么茶树在低温环境下会受到哪些影响呢?

一、茶树常见的冻害类型
茶树常见的冻害有雪冻、霜冻及干冷风冻几种。长江以南产茶区以雪冻和霜冻为主,长江以北产茶区三种冻害均有发生。茶树受冻后有赤枯和青枯两种表现形式,长江以南以赤枯状为常见,长江以北赤枯、青枯兼有发生。

二、茶树冻害的发展顺序
茶树具有一定抵御低温的能力,但不同的茶树器官耐寒力不同。就叶、茎、根各器官而言,其耐寒能力依次递减。但是在露地栽培条件下,由于叶对茎、茎叶对根来说,具有保护作用,因此在越冬期间,茶树树冠上部、中部和根部的温度,总是自上而下依次递增,所以在生产实践中,茶树冻害的发展顺序,往往表现为顶部枝叶首先受害,进而波及茎部,只有在极度严寒的条件下,根部才受害而致全株死亡。

三、低温导致茶树冻害的机理
低温导致茶树冻害,其机理是叶片组织内部细胞结冰。当温度降至—1.07 ℃时,首先是细胞间隙自由水开始结冰,形成冰的核心。温度继续下降则引起冰体不断扩大,吸取原生质体中的水分,导致原生质缓慢失水变性;同时冰晶体由小变大,对细胞产生挤压性机械损伤;由于原生质与细胞壁对水分反应速度有差异,温度骤升骤降时,细胞内产生质壁撕扯和分离,细胞会损伤;在温度迅速回升时又产生水分胁迫,细胞间隙水快速蒸发,原生质吸水补给不及时导致失水变性。当温度急剧下降,异常寒冷时,还会使细胞结冰,损伤细胞膜,导致细胞死亡。

上世纪80年代的研究表明,茶树霜冻害的发生与茶树叶面的冰核细菌种类和数量有密切关系。茶树叶表有了冰核细菌,通常以冰核细菌作为冰的核心开始形成冰体,通常在—2 ~—5 ℃,冰核细菌会诱发植物细胞水结冰而发生霜冻,无冰核细菌存在的茶树叶片,可耐受—7 ~—8 ℃低温而不发生霜冻。茶树上主要的冰核细菌种类有菠萝泛菌(Pan talananatis)、成团泛菌(Pantoea aggloinerans)、甘兰黄草胞菌(Xanthomonas campetris)、菠萝果腐欧文氏菌(Erwinia ananas)等,因此对茶树叶面冰核细菌的防治有助于减轻霜冻害的发生。
四、影响茶树冻害的因素
1. 气象条件与茶树冻害的关系
当月平均气温、极端最低气温、负积温、连续低温天数与冻害密切相关,其绝对数值越大,冻害越重,反之冻害较轻或不受冻。
2. 冻土深度与茶树冻害的关系
茶树冻害程度的轻重与地温极值、负积温、最大冻土深度、连续冻土日期呈正相关,如茶树长期处于冻土层中,会造成茶树根系对水分吸收转运困难,以至地上部脱水枯死,根系萎缩腐朽。
3. 大气相对湿度与茶树冻害的关系
凡是冬季雨水偏少,大气相对湿度偏低,土壤又缺水的情况下,茶树冻害往往较重,若出现大风,短期内茶树即会出现青枯型冻害。在我国华南茶区,这类茶树冻害不多见,而在江北茶区常有发生。
4. 茶树冻害与茶树品种、树龄的关系
不同茶树品种的耐寒力有差异,如云南大叶茶由于长期生长在冬无严寒、夏无酷暑的环境中,相比其他品种,它的耐寒能力较弱,通常在气温为0.5 ℃时,就可能发生冻害。而一般中小叶种的茶树,耐寒能力比云南大叶茶强,在低温持续时间不长的情况下,能耐—10 ℃低温。
茶树受冻程度的轻重,与树龄的大小也有一定的关系。表现为随树龄的增加,耐寒能力相应增强。
5. 茶树冻害与地理条件的关系
随海拔和纬度的增高,越冬期的绝对低温、负积温总值、低温持续天数逐渐增高。所以,在高纬度、高海拔的立地条件下,茶树容易受冻。
“雪打高山,霜打洼”是一句气象农谚。高山降雪量多于平地和丘陵洼地,由于冷空气下沉,冬季常出现浓霜。在这些地理位置上种茶,茶树极易产生雪冻和霜冻。此外,风口和冷空气过道也易出现冻害,受害茶树会呈带状分布。
6. 茶园管理与茶树冻害的关系
茶园管理技术运用得当,可增强茶树长势,提高茶树耐寒能力,达到茶树安全越冬和减轻茶树冻害程度的作用;反之,如管理不当,将会加重茶树冻害的发生和发展。
有关茶树低温冻害的预防和补救措施会在后续文章中发布,敬请关注~
本文节选自《中国茶经》,陈宗懋、杨亚军主编。部分图片来源于网络。
来源:中国茶叶
如涉及版权问题请联系删除
茶树是重要的经济作物,起源于中国并在世界范围内产生重要影响。令人愉悦的香味和沉淀千年的文化使茶成为除水之外最受大众喜爱的健康饮品。近年来随着测序技术的发展,多个茶树全基因组物理图谱被破译[1-8],但仍然缺乏对乌龙茶之首铁观音基因组的研究。
由福建农林大学尤民生教授和中国农业科学院(深圳)农业基因组研究所(以下简称“基因组所”)张兴坦研究员主持,联合国内外多家单位的科学家共同合作成功破解铁观音基因组和茶树演化史,研究成果于2021年7月15日以“Haplotype-resolvedgenomeassemblyprovidesinsightsintoevolutionaryhistoryoftheteaplantCamelliasinensis”为题发表在国际顶级期刊《自然遗传学》(NatureGenetics)上。研究人员利用独立开发的新算法攻克高杂合、高重复铁观音基因组组装难题,并在此基础上阐述了等位基因不平衡、茶树群体演化和驯化等相关科学问题。
成功破解铁观音单倍体分型组装技术难题
铁观音原产于福建省泉州市安溪县西平镇,相传于清朝雍正年间被当地茶农发现,因其叶形似观音脸重如铁而被乾隆赐名为“铁观音”。
茶树是自交不亲和植物,更因长期的无性繁殖积累大量体细胞突变,导致基因组高度杂合、组装难度很大。
研究团队利用自主开发的两种算法(Khaper和ALLHiC)整合Illumina短读长、PacBioCLR长读长和高通量三维染色质捕获技术(Hi-C),攻克高杂合基因组组装难题,成功拼接了两套铁观音基因组(图1):单倍体参考基因组(monoploidreferencegenome)和单倍体分型基因组(haplotype-resolvedgenome)。茶树是二倍体,含有15对同源染色体(2n=2x=30),单倍体参考基因组是筛选同源染色体中的一份拷贝作为代表组装到染色体水平,而单倍体分型基因组是将来源于不同父母本的两套同源染色体同时组装到染色体水平。前者不区分等位基因,广泛用于二倍体基因组的组装;后者区分等位变异,更完整地呈现二倍体基因组的全部遗传信息。

图1.铁观音基因组组装和质量评估。(a)单倍体参考基因组circos图,呈现15染色体特征;(b)Hi-C热图呈现15条染色体组装质量;(c)LAI评估铁观音和已发表其他茶树基因组组装质量;(d)铁观音单倍体参考基因组和分型基因组的共线性比较。
利用等位优势基因应对“遗传负荷”
铁观音距今已有约300年的栽培历史,长期的无性繁殖积累大量体细胞突变(包括有害突变),增加了遗传负荷(geneticload),导致其适应性降低。然而人们对无性繁殖作物如何应对遗传负荷这一问题知之甚少。
传统的杂种优势现象可以由显性效应和超显性效应两种假说解释:显性效应指个体倾向于利用有利于生长和发育的优势等位基因(或显性基因)而忽略对个体不利的劣势基因(或隐形基因);超显性效应指杂合等位组合在多种生境下优于任一纯合等位的现象。
研究人员基于铁观音分型基因组组装,鉴定到14691个基因具有等位变异。RNA-seq分析显示,其中1528个基因存在一致性的等位特异性表达(consistentallele-specificexpression,consistentASE),即其中一个等位基因在所有组织和样本中的表达都高于另一等位基因;而只有386个基因存在非一致性的等位特异表达(inconsistentASE),即两个等位基因分别在不同的组织中存在特异高表达。前者可以被认为是具有显性效应的基因,而后者是具有超显性效应的基因。这一结果显示,在无性繁殖的茶树基因组中,显性效应可能是其应对遗传负荷的重要机制。面对大量积累的体细胞突变或有害突变,个体选择使用未突变或对个体有利的等位基因维持其正常的生长发育和对环境的适应性。
大叶茶与小叶茶存在不同的演化和驯化历史
茶树遗传多样性较高。
研究人员对161个茶树品种和15个近缘种大理茶进行重测序分析发现,这些个体聚类为三组,分别为大理茶(CT)、大叶茶(CSA)和小叶茶(CSS)。其中大叶茶分为两个亚组,古大叶茶(ACSA)和栽培大叶茶(CCSA);而小叶茶分为四个亚组,依据其主要地理分布可以划分为川陕赣(SSJ)、浙江和闽北(ZJNFJ)、闽南(SFJ)、两湖(湖南和湖北)和安徽(HHA)。
遗传分析显示各茶区存在频繁的种质基因交流,其中一些与有记录的茶树杂交育种历史相吻合(图2)。比如茶树黄玫瑰品种呈现出铁观音与黄棪基因组混合的组分,而黄玫瑰是两者子代黄观音和黄棪回交选育的优良品种。这种频繁的基因交流不仅出现在茶树种内,在茶树与近缘种间也普遍存在。研究人员通过对8个茶树品种和山茶属茶组的12个近缘物种系统发育分析发现,这些物种呈现网状演化模式,而非简单的树形演化。更多的证据表明,茶树与近缘种间频繁的杂交渐渗(introgression)是其网状演化和维持茶树遗传多样性的重要因素。
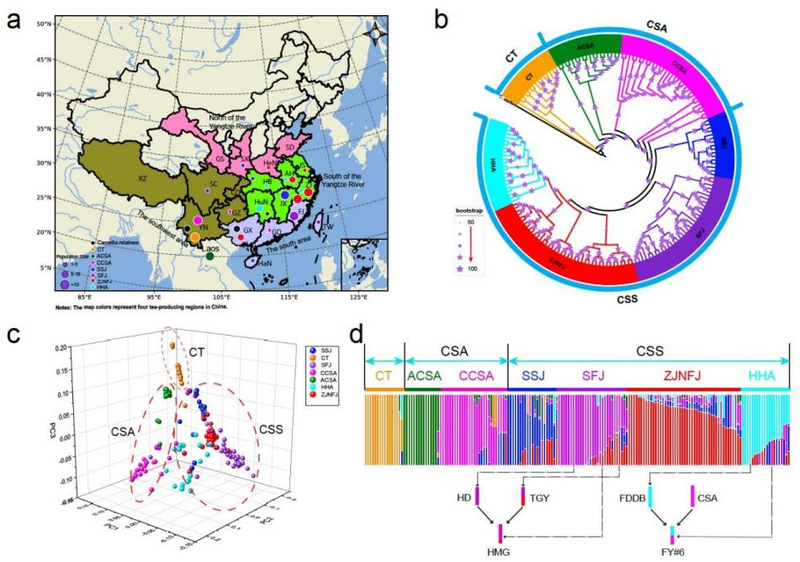
图2.茶树的系统发育关系和群体结构。(a)重测序个体的地理分布;(b)系统发育树;(c)重测序群体的主成分分析(PCA);(d)K=7模型下的群体遗传结构图,下方显示了两个有记录的现代繁殖事件。
栽培茶树品种主要分为大叶茶(C.sinensisvar.assamica,CSA)和小叶茶(C.sinensisvar.sinensis,CSS),两者之间在形态上具有明显差异。前者植株较高、叶片较大、多为乔木,主要生长在云南、广东等温度较高的地域;而后者植株较矮、叶片较小、多为小乔木或灌木,可以生长在北至山东等较寒冷地区。
研究人员通过群体遗传分析发现大叶茶和小叶茶具有不同的演化和驯化历史。在距离约259-181万年的格拉斯阶时期,剧烈的气候变化很可能导致了整个茶树物种的群体收缩,这也是一次大叶茶和小叶茶共享的瓶颈事件。两个变种分化后,小叶茶的生境遭遇了末次冰盛期,2.65-1.9万年前的温度骤降可能使得小叶茶出现了再一次的群体瓶颈,但随后适应了环境的小叶茶迅速扩张,群体规模得到恢复(图3)。

图3.茶树的群体动态历史。(a)小叶种(上)和大叶种(下)历史有效群体大小;(b)过去42万年的历史温度变化。
此外,人们对大叶茶和小叶茶制品的偏爱有所不同也导致了两者经历了平行的驯化历程(图4)。大叶茶早期的驯化主要筛选了一些糖苷类物质转运的相关基因,而在品种改良阶段人们更关注生物碱和香气挥发物相关的代谢途径。
例如,研究人员鉴定到CsXDH基因在大叶茶品种改良阶段受到强烈的人工选择,该基因编码黄嘌呤脱氢酶,是咖啡因合成通路的重要基因[9]。
小叶茶早期人工选择的基因与植物防御和抗性相关,事实上这些受选择的基因同时也参与到了重要的次级代谢产物的生物合成,例如R-柠檬烯、β罗勒烯等途径。
作为儿茶素合成通路的关键基因,F3’H也在这一过程中受到人工选择。而在小叶茶品种改良过程中,人工驯化涉及到花发育和一氧化氮(NO)响应相关的基因。之前的研究表明,NO的积累可以加速γ-氨基丁酸的消耗从而帮助植物抵御冷胁迫[10],这一结果暗示着筛选耐寒的品种也是人工选育的重要目标。
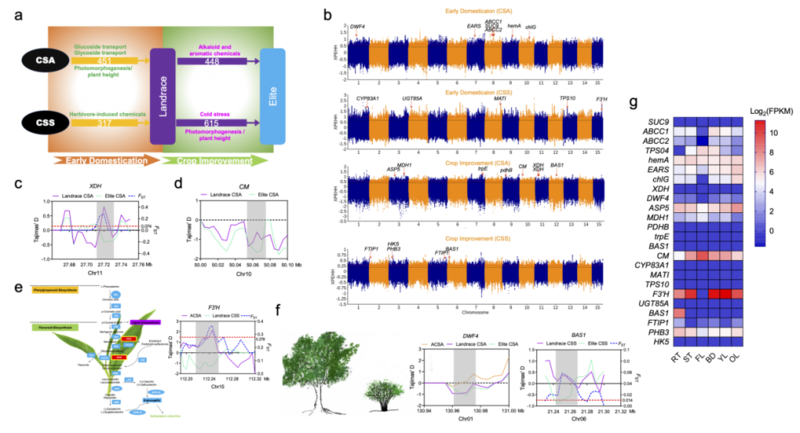
图4.大叶茶(CSA)和小叶茶(CSS)人工选择的特征与平行驯化的证据。(a)CSA与CSS平行驯化路线图;(b)基于XP-EHH识别的选择性清除信号在全基因组的分布;(c)XDH基因中人工选择的信号;(d)CM(脉络膜类诱变酶)基因中的人工选择信号;(e)F3’H基因中人工选择的信号;(f)与植物高度相关的BAS1和DWF4基因的人工选择信号;(g)人工选择基因的表达,包括根(RT)、茎(ST)、花(FL)、芽(BD)、幼叶(YL)、老叶(OL)。
茶树的“绿色革命”基因
20世纪60年代,大规模推广矮秆或半矮秆的水稻和小麦品种极大的提高了作物产量,解决了发展中国家急剧增长的粮食需求,控制水稻株高的sd1基因和小麦的rht基因也因其巨大的贡献被称为“绿色革命基因”[11]。研究人员发现,茶树的株高在长期的栽培过程中也受到驯化,体现在两个细胞色素P450家族基因受到人工选择(CsBAS1和CsDWF4)。这两个基因参与油菜素内酯合成,前者的拟南芥突变体导致了延长的下胚轴,而后者的突变直接导致了植株侏儒的表型[12-13]。
该项目在攻克铁观音基因组的基础上,通过对茶树种群水平的遗传分析,揭示了该物种的演化和人工驯化历史。该成果为利用组学分析和分子生物学技术挖掘功能基因、解析其背后的遗传调控机制,开展基于大数据驱动的基因组智能设计育种奠定了坚实的理论基础,同时也为缩短育种周期、提高育种效率、降低育种成本提供了科学依据。
该项目由福建农林大学、基因组所等国内外多家单位合作完成。基因组所张兴坦研究员、福建农林大学硕士研究生陈帅、福建省农科院水稻所施龙清博士、中国农科院烟草研究所龚达平研究员为论文共同第一作者,福建农林大学尤民生教授、唐海宝教授和基因组所张兴坦研究员为共同通讯作者。
来源:农业基因组所报道,信息贵在分享,如涉及版权问题请联系删除


































